
In brief:
In our recent study in Translational Psychiatry with first authors Jason He and Georg Oeltzschner show that difficulties with sensory processing at the perceptual and clinical level in children autism are associated with neurochemical differences in the brain regions responsible to sensory processing
Some background
Sensory differences experienced by people on the autism spectrum are well-known. Sometimes, these sensory differences can be strengths and indeed, some autistic individuals are better able to detect and discriminate sensory stimuli than neurotypical individuals. Still, many individuals on the autism spectrum state that their sensory differences can be incredibly debilitating; for example the brightness of lights, certain smells or tastes, and certain kinds of touch. Increased evidence suggests that sensory difficulties (which emerge early in life) might exacerbate some of the social core difficulties in autism.
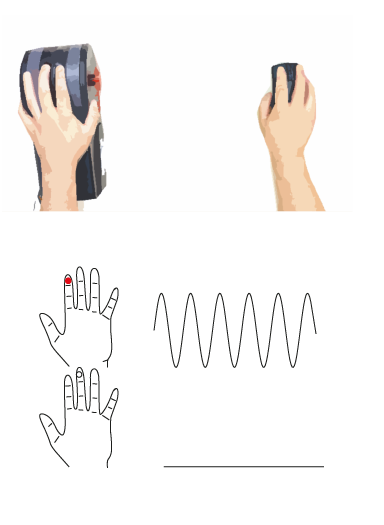
Strong responses to sensory input are referred to as hyperreactivity, while the absence of an expected response is referred to as hyporeactivity. However, these are often measured through questionnaires. We are interested in studying how differences in sensory sensitivity (i.e. how well can we perceive simple tactile stimuli, or differences between stimuli). We use a technique called psychophysics to measure this in children (click here for more information) and have shown differences in sensory perception in children with autism and ADHD (click here for more information).
One of the theories that we believe to explain sensory differences in autism is the theory of ‘altered excitation-inhibition balance’. Broadly, the altered excitation-inhibition balance theory posits that sensory difficulties in autism are due to an imbalance in the neurochemicals responsible for excitatory and inhibitory signalling. Excitatory signalling is principally dependent on glutamate, while inhibitory signalling is principally dependent on gamma-aminobutyric acid (better known as ‘GABA’). Evidence for this theory comes from a variety of sources; This includes animal models of autism that have shown alterations of the genes that encode for GABA and glutamate in autism and have linked these alterations to sensory hyperreactive behaviours in mice, genetic and anatomical data, and others.
Our current study

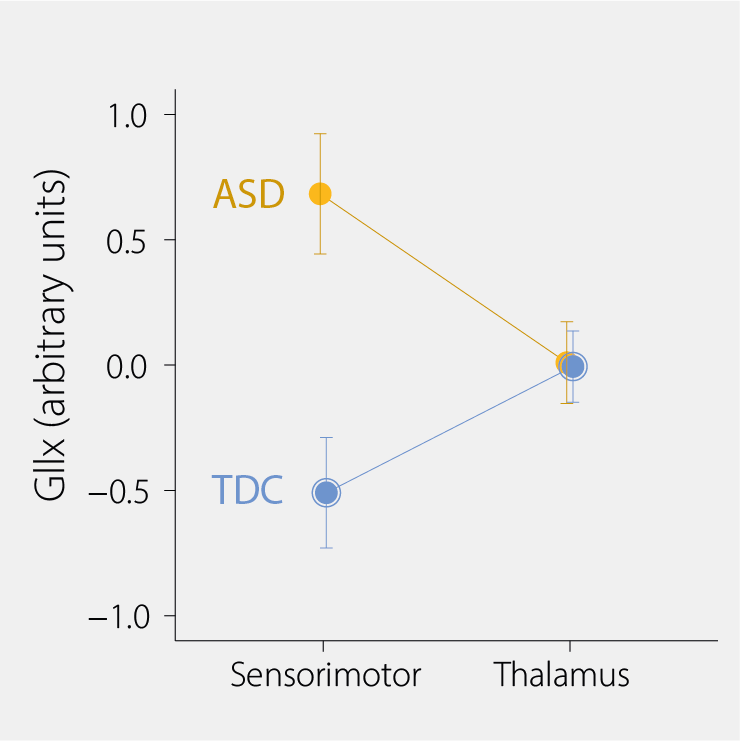
In our recently published study in Translational Psychiatry, we measured both sensory sensitivity and sensory reactivity, and used a technique called magnetic resonance spectroscopy (MRS) to measure the amount of Glx (glutamate+glutamine) and GABA in the brains of autistic children and neurotypical controls.
We measured these metabolites in the sensorimotor cortex and in the thalamus, both regions having strong involvement in sensory processing. While we’re measuring the same Glutamate and GABA that are responsible for neurotransmission, it should be noted that we measure all of the Glx and GABA, and thus we are likely measuring GABA/Glx metabolism rather than neurotransmission. Glx is a compound measure of Glutamate and Glutamine.
Unlike previous work, we did not reproduce the finding that autistic children had different levels of GABA in the areas of the brain responsible for sensory processing. What we did find, however, was elevated levels of glutamate in the sensorimotor cortex. Although we can’t say this is directly proportional to increased excitatory neurotransmission, increased glutamate does support the excitation-inhibition imbalance theory, suggesting that, on average, individuals on the autism spectrum have increased metabolism related to excitation than that contributing to ‘inhibition’ (in these specific parts of their brains) than neurotypicals.
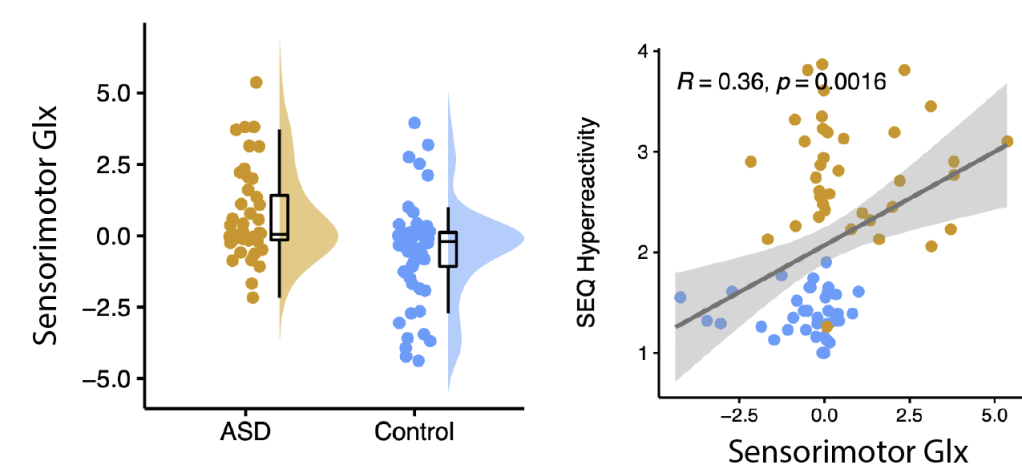
We also explored the associations between brain neurometabolites and sensory differences in autism. We found that in addition to having more glutamate in primary sensorimotor, these glutamate levels were predictive of both parent-reported sensory hyperreactivity and sensory sensitivity to touch in autistic children. Specifically, more glutamate in the sensorimotor cortex was associated with more hyper and hyporeactivity. More glutamate in the sensorimotor cortex and thalamus was also associated with more difficulties discriminating between tactile (i.e., touch) stimuli. These findings further support the excitation-inhibition imbalance theory of sensory symptoms in autism.
Finally, our data also suggest that the relationship between sensorimotor and thalamic glutamate is different in those with autism, suggesting a hyperglutamatergic connectivity (possibly suggesting increased thalamocortical connectivity).
Clearly, more investigation is needed. However, our results show that sensory differences in autism may indeed be explained by alterations of excitation-inhibition balance in the brain.
But of course there are limitations
While our results are exciting in that they support a long-held theory, we need to be careful about how we interpret these results as mentioned above. MRS measures of glutamate and GABA only provide measures of total concentration (i.e., the amount of glutamate and GABA) in certain areas of the brain. Concentrations do not tell us how about the functioning of these neurotransmitters in the areas of the brain we investigated. Although we can assume that more glutamate or less GABA means more excitatory signalling and less inhibitory signalling, this is merely an assumption.
Moreover, our study only assessed glutamate and GABA levels in high functioning autistic children that were predominantly male, and we are currently unsure how well our results would translate to those on the spectrum that were not well represented in our sample (e.g., autistic females or non-verbal autistic children).
